
Even in the frenzy of the blood and the pain, the snarling and dying at a kill site there is a choreography that has evolved to share the spoils and reduce conflict. The patterns that predators leave behind often provide clues pointing to their identity–even if the site is 12,000 years old and absent distinctive tooth marks. Gary Hanes has spent years studying the kill sites of various predators and his research provides a general picture of their different patterns.
According to Haynes (1988) every wolf pack has its hierarchy and once the prey is dead, and often even before, the process of staking out claims on preferred cuts and dividing the carcass begins. The dominant wolf will take the choice position. The blood and internal organs inside the abdomen are a favorite first pick. Sternal elements and ribs are usually damaged in the process. Other high ranking wolves will claim the rump and upper legs where there are large masses of flesh. If there are more animals in the pack than can comfortably situate themselves around the carcass to eat without invading each other’s space, they’ll start disarticulating the limbs, causing distinctive damage on the proximal ends of the femora and humeri , and their anchoring points on the pelvis and scapulae. The prizes will be carried a short distance away to be gnawed on in relative solitude. Lower ranking wolves will tear off smaller less desirable parts (e.g. ears, tail, jaw/tongue) and carry them further away. These satellite consumption spots will be randomly distributed around the carcass and about 20 feet apart (Haynes, ibid.). Tooth marks and distance will be correlated as wolves lower in the pecking order tend to invest more time gnawing on their meager rations instead of trying to muscle in and steal another portion.
The scatter of bones at a wolf kill site rarely exceeds 100 ft. from ground zero. Within this area one can expect to find the skull, ribs and vertebrae, but rarely the sternal elements, patelae (knee bones) and caudal (tail) vertebrae. A few of the major limb bones are likely to be missing—more if scavenging has been heavy. The lower legs of hoofed animals offer more tendons and ligaments than muscle so they are often discarded uneaten and the bones are often found in anatomical sequence. If scavenging is heavy, the lower leg may represent a prized morsel and elements may be transported miles from the kill site (Haynes, 1985). The scatter is normally lower at a non-kill/scavenging site, though that will increase over time with scavenging by a host of creatures and kicking. Most, if not all of the major limb bones should be close by. Wolves will normally leave even a fresh-dead prey carcass relatively intact. Unless they are particularly hungry, wolves simply prefer to do their own “shopping” (Haynes, 1982).
Wolves follow predictable sequences in disarticulating and consuming a kill. Their pattern of utilization varies with the prey species and its particular anatomical characteristics, its size, how hungry the wolves are, how many individuals are in the pack, and the season. The chart below summarizes his findings for large prey over 300 kg (~660 lbs.–e.g. moose or bison size). According to Haynes, the patterns are so regular for a particular predator and prey species that out-of-sequence disarticulation or damage, or the absence of a normal step, are reliable indicators of scavenger activity instead of predation, or other disturbance processes such as trampling, etc. (Haynes, 1982).
Carnivore Utilization Stages |
|||
|
|
Stage I: light to moderate |
Stage II: Full |
Stage III: Heavy |
|
sternal ends consumed |
Many on one side broken off below their articulating ends and scattered |
further broken up |
|
|
greater trochanter damaged & trochlear rim scored |
disarticulated from pelvis, distal condyles gouged. Toothmarks undercut head. shaft lightly scored |
head removed, distal end gone, shaft broken |
|
|
Tibia |
|
still articulated to femur |
disarticulated, proximal end gone |
|
Humerus |
head and greater tuberosity furrowed and gouged |
disarticulated from scapulae, greater tuberosity gone, shaft lightly scored |
proximal end gnawed off, approx. 1/3 of proximal shaft gone. |
|
Ulna |
olecranon process damaged |
|
|
|
edges of ilia and ischia gnawed |
|
gnawed down to acetabula |
|
|
vertebral border damaged, still attached to humerus. |
disarticulated from humerus, edge splintered |
gnawed down to glenoid process |
|
|
vertebrae |
|
spines and lateral processes gnawed or broken off |
Only a few badly gnawed vertebrae remain articulated |
|
Skull |
Still articulated to the body, no damage to bones. Nasal cartilage gnawed |
disarticulatedNasal bones gnawed |
Only toothrows remain from head
|
|
partly defleshed, articulated |
defleshed. |
Mandibles disarticulated |
|
adapted from Haynes1982 and 1999
It’s probably too soon to use scatter diagrams and limb bone tallies to test the predator theory with the sloths. We still have the entire south bank of the creek to explore and map. However, we do have a large sample of bones to appraise using Haynes’s rules of carcass utilization. The ribs we have range from perfect to stage III. Our sole femur (R), the truest indicator of predation according Haynes (1982), shows signs of light scavenging by small mammals but none of the damage to the greater trochanter that he predicts from disarticulation by predators. The other femur (L) is missing entirely. Both tibiae are missing as well. Humeri heads are toothmarked and overall the bone shows Stage III damage, as do the vertebrae and the pelvis. The one ulna that survived (R) looks untouched—no signs of the disarticulation damage at the olecranon process (elbow) that he predicts in the case of a kill. One scapula (R) is almost pristine, but the other one is heavily damaged by trampling–no evidence of gnawing however. The skull is stage II while the mandibles are stage III.
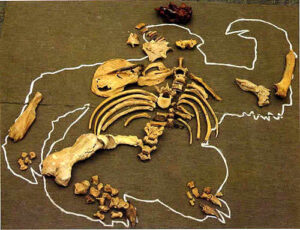 Conclusion: no traces of disarticulation by predators judging from the adult bones we’ve found so far. However, most of the “baby’s” bones are still missing. All of the “toddlers” major bones are AWOL too, except for the distal half of one humerus. If we assume the baby weighed about 90# and the toddler perhaps 300# (based on scapulae, the only common bone we have), that’s about 200# of flesh–a healthy dinner for a pack of wolves. How much would a pack of wolves eat at one sitting? Would they have turned their attention to the tender youngsters before eating a tough old adult? Would they have abandoned the adult’s carcass only half eaten? What accounts for the disparity in utilization stages of the carcass and why are the bones from the right side better preserved than those from the left? Hayne’s rules would say even if predation is confirmed, the deviations from the expected pattern indicate there are other factors at work. . . but he also warns that the rules may be entirely different for ground sloths. As ever, we have more questions than answers, but Haynes’s research holds out the promise that with some more data maybe we can start to peel back the layers. . . . Dave
Conclusion: no traces of disarticulation by predators judging from the adult bones we’ve found so far. However, most of the “baby’s” bones are still missing. All of the “toddlers” major bones are AWOL too, except for the distal half of one humerus. If we assume the baby weighed about 90# and the toddler perhaps 300# (based on scapulae, the only common bone we have), that’s about 200# of flesh–a healthy dinner for a pack of wolves. How much would a pack of wolves eat at one sitting? Would they have turned their attention to the tender youngsters before eating a tough old adult? Would they have abandoned the adult’s carcass only half eaten? What accounts for the disparity in utilization stages of the carcass and why are the bones from the right side better preserved than those from the left? Hayne’s rules would say even if predation is confirmed, the deviations from the expected pattern indicate there are other factors at work. . . but he also warns that the rules may be entirely different for ground sloths. As ever, we have more questions than answers, but Haynes’s research holds out the promise that with some more data maybe we can start to peel back the layers. . . . Dave
References
Haynes, G. 1982. Utilization and skeletal disturbances of North America prey carcasses. Arctic 35: 266-281.
Haynes, G. 1985. On watering holes, mineral licks, death, and predation. In Environments and Extinctions: Man in late glacial North America, Eds. J Mead and D Mettzer. Center for the Study of Early Man.
Haynes G. 1988. Prey bones and predators: potential information from analysis of bone sites. Ossa: 7: 75-97.
Haynes, G. 1999. The role of mammoths in rapid Clovis dispersal. In Mammoth and mammoth fauna: studies on an extinct ecosystem. Proceedings of the first International Mammoth Conference, St. Petersburg, Russsia. P. 9-38.